Search
{{#data.error.root_cause}}
{{/data.error}}
{{^data.error}}
{{#texts.summary}}
[{{{type}}}] {{{reason}}}
{{/data.error.root_cause}}{{texts.summary}} {{#options.result.rssIcon}} RSS {{/options.result.rssIcon}}
{{/texts.summary}} {{#data.hits.hits}}
{{#_source.featured}}
FEATURED
{{/_source.featured}}
{{#_source.showImage}}
{{#_source.image}}
 {{/_source.image}}
{{/_source.showImage}}
{{/_source.image}}
{{/_source.showImage}}
{{/data.hits.hits}}
{{{_source.title}}} {{#_source.showPrice}} {{{_source.displayPrice}}} {{/_source.showPrice}}
{{#_source.showLink}} {{/_source.showLink}} {{#_source.showDate}}{{{_source.displayDate}}}
{{/_source.showDate}}{{{_source.description}}}
{{#_source.additionalInfo}}{{#_source.additionalFields}} {{#title}} {{{label}}}: {{{title}}} {{/title}} {{/_source.additionalFields}}
{{/_source.additionalInfo}}Anti-Lhcb2 | LHCII type II chlorophyll a/b-binding protein
(Cat#: AS01 003)
Description
- Immunogen: BSA-conjugated synthetic peptide derived from a highly conserved sequence of Lhcb2 proteins from angiosperms (monocots and dicots) and gymnosperms, including Arabidopsis thaliana Lhcb2.1 UniProt: Q9SHR7, TAIR: AT2G05100, Lhcb2.2 UniProt: Q9S7J7, TAIR:AT2G05070, Lhcb2.3 UniProt:Q9XF87, TAIR:AT3G27690
- Host: Rabbit
- Clonality: Polyclonal
- Purity: Immunogen affinity purified serum in PBS pH 7.4.
- Format: Lyophilized
- Quantity: 50 µg
- Reconstitution: For reconstitution add 50 µl of sterile water
- Storage: Store lyophilized/reconstituted at -20°C; once reconstituted make aliquots to avoid repeated freeze-thaw cycles. Please remember to spin the tubes briefly prior to opening them to avoid any losses that might occur from material adhering to the cap or sides of the tube.
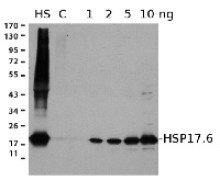
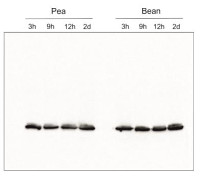
- Tested applications: Immunoprecipitation (IP), ImmunoGold (IG), Western blot (WB)
- Recommended dilutions: 5 µl of antibody solution (IP), 1: 100 (IG), 1: 500 - 1 : 5000 (WB)
- Expected | apparent MW: 28.6 | 25 kDa for Arabidopsis thaliana
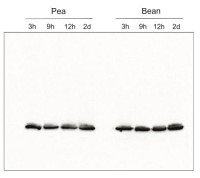
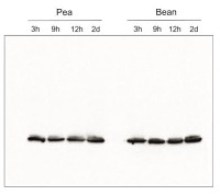
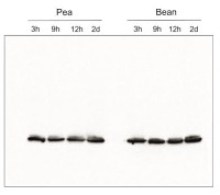
- Confirmed reactivity: Acer pseudoplatanus, Arabidopsis thaliana, Arachis hypogaea, Brachypodium sylvaticum, Camelina sinensis, Cicer arietum, Chlorella vulgaris, Colobanthus quitensis Kunt Bartl, Chlamydomonas reinhardtii, Cucumis sativus, Cytisus cantabricus (Wilk.) Rchb. F., Hieracium pilosella L., Hieracium pilosella L., Hordeum vulgare, Lasallia hispanica, Lycopersicon esculentum (Solanum lycopersicon), Miscanthus x giganteus, Mesembryanthemum crystallinum, Nicotiana benthamiana, Nicotiana tabacum, Oryza sativa, Pisum sativum, Phaseolus coccineus L., Phaseolus vulgaris, Physcomitrium patens, Setaria viridis, Sinapsis alba, Spinacia oleracea, Syntrichia muralis (Hedw.) Raab, Triticum aestivum, Triticale, Zea mays
- Not reactive in: No confirmed exceptions from predicted reactivity are currently known
- The major light-harvesting antenna complex II (LHCII) in photosynthetic eukaryotes is located in the thylakoid membrane of the chloroplast. It is a heterotrimeric complex formed by up to 3 different individual subtypes of chlorophyll a/b-binding proteins: Lhcb1, Lhcb2, and Lhcb3. Lhcb2 is often coded by several nuclear genes and is found together with Lhcb1 within the mobile LHCII trimers involved in state1-state2 transition.A molecular characterisation of the LHCII proteins can be found in Caffarri et al. (2004) A Look within LHCII: Differential Analysis of the Lhcb1−3 Complexes Building the Major Trimeric Antenna Complex of Higher-Plant Photosynthesis. Biochemistry 43 (29): 9467–9476.
- Ermakova et al. (2024). Chloroplast NADH dehydrogenase-like complex-mediated cyclic electron flow is the main electron transport route in C4 bundle sheath cells. New Phytol. 2024 Jul 22.doi: 10.1111/nph.19982.Zhao et al. (2024). Psb28 protein is indispensable for stable accumulation of PSII core complexes in Arabidopsis.Plant J. 2024 May 26. doi: 10.1111/tpj.16844. Ciesielska et al. (2024). S2P2-the chloroplast-located intramembrane protease and its impact on the stoichiometry and functioning of the photosynthetic apparatus of A. thaliana. Front Plant Sci. 2024 Mar 15:15:1372318. doi: 10.3389/fpls.2024.1372318. Kim et al. (2024).Photoautotrophic cultivation of a Chlamydomonas reinhardtii mutant with zeaxanthin as the sole xanthophyll. Biotechnol Biofuels Bioprod. 2024 Mar 14;17(1):41. doi: 10.1186/s13068-024-02483-8. Ye et al. (2023). The light-harvesting chlorophyll a/b-binding proteins of photosystem II family members are responsible for temperature sensitivity and leaf color phenotype in albino tea plant. J Adv Res . 2023 Dec 25:S2090-1232(23)00404-6.doi: 10.1016/j.jare.2023.12.017. Singh, Muthamilarasan, Prasad (2022). SiHSFA2e regulated expression of SisHSP21.9 maintains chloroplast proteome integrity under high temperature stress. Cell Mol Life Sci. 2022;79(11):580. Published 2022 Nov 3. doi:10.1007/s00018-022-04611-10Cazzaniga et al. (2022). Engineering astaxanthin accumulation reduces photoinhibition and increases biomass productivity under high light in Chlamydomonas reinhardtii. Biotechnol Biofuels Bioprod. 2022 Jul 11;15(1):77. doi: 10.1186/s13068-022-02173-3. PMID: 35820961; PMCID: PMC9277849.Bru, Steen, Park, et al. (2022) The major trimeric antenna complexes serve as a site for qH-energy dissipation in plants. J Biol Chem. 2022;298(11):102519. doi:10.1016/j.jbc.2022.102521Ivanov et al. (2022) The decreased PG content of pgp1 inhibits PSI photochemistry and limits reaction center and light-harvesting polypeptide accumulation in response to cold acclimation. Planta 255, 36 (2022). https://doi.org/10.1007/s00425-022-03819-0Bychkov et al. (2022) The role of PAP4/FSD3 and PAP9/FSD2 in heat stress responses of chloroplast genes. Plant Sci. 2022 Sep;322:111359. doi: 10.1016/j.plantsci.2022.111359. Epub 2022 Jun 20. PMID: 35738478.Lang et al. (2011).Simultaneous isolation of pure and intact chloroplasts and mitochondria from moss as the basis for sub-cellular proteomics. Plant Cell Rep. 2011 Feb;30(2):205-15.doi: 10.1007/s00299-010-0935-4.
- Immunoprecipitation has been done using Immunoprecipitation kit from Roche, Cat.No. 11 719 386 001.Protein is processed into mature form (Jansson 1999).
Boca Scientific is your premiere source for high-quality, innovative solutions for Cell Biology, Molecular Biology, Immunology, genetics and other lab products and reagents. We bring leading-edge products from our own-line and around the world to laboratories in the US and Canada. Our goal is to offer excellent solutions to drive research and discoveries backed by superior customer support.

{kind=link}
{kind=link}